11. 1. 3. La notion d’organisme
Cet ensemble solidaire dont les éléments sont en synergie se voit attribuer le nom d’organisme par Humboldt371.
Ce terme est emblématique de la période, tant au plan lexicologique, épistémologique, que métaphorique. Il apparaît en français en 1729 où il désigne alors tout être vivant doté d’organes. C’est en 1802 qu’il désigne l’ensemble des organes qui constituent un être vivant. En 1842, il s’adjoint le sens figuré de « ensemble composé d’éléments structurés » (en politique, en économie). Sous l’influence du français, il apparaît durant le 18e siècle allemand au sens de « être vivant, organisme autonome, ensemble structuré, unitaire et articulé ». En anglais, le terme apparaît en 1664 au sens de « structure organique ». En 1768, il prend le sens figuré de « système organisé », de « tout formé de parties interdépendantes ». C’est en 1842 qu’il adopte sa définition actuelle de « corps organisé, consistant en un ensemble de parties interconnectées et dépendantes, et dont la communauté vise au maintien de la vie ». En d’autres termes, un organisme désigne la structure matérielle d’un animal ou d’une plante. L’affinement du concept va clairement vers le sens d’interconnexion organisatrice à visée structurante372.
Le concept d’organisme est à relier à cet animisme universel qu’est la théorie des monades de Leibniz. Par l’analyse de l’être organisé qu’il pratique dans la Critique du jugement (1790), Kant contribue à le faire entrer parmi les concepts phare de la biologie et de la philosophie romantique. D’usage fréquent chez Humboldt et Bopp, il sera employé par Comte (microcosme organique), qui demeure influencé par la pensée romantique, et deviendra un modèle d’économie politique chez Claude Bernard. Envisagé comme un élément spéculatif assurant un rôle transitionnel entre la nature et l’esprit, il ne doit pas être réduit à une simple métaphore explicative ou poétique. Pour le monde romantique, l’organisme représente un principe explicatif universel (cf. Ernst Cassirer (1972 : 101)).
L’ouvrage de Friedrich von Schlegel, Ueber die Sprache und Weisheit des Indier (1808), inaugure l’utilisation de ce concept dans l’étude des langues. OEuvre transitionnelle qui, comme l’indique son titre, mêle philosophie et théorie du langage, elle introduit une nouvelle discipline et un terme dans le champ de l’étude des langues : la grammaire comparée (vergleichende Grammatik)373. Schlegel envisage un classement de langues par comparaison de leurs types grammaticaux et aboutit à une classification bipartite opposant les langues organiques (i. e. flexionnelles), « ‘fondées sur la puissance et la fécondité de leurs racines ’», et les langues mécaniques (i. e. isolantes) « ‘produits d’aménagements compliqués et fragiles’ »374. En 1818, dans ses Observations sur la langue et la littérature provençale (écrites en français), son frère August Wilhelm adopte une perspective historiciste et ajoute une classe à la bipartition de son cadet. Les langues sont désormais divisées en langues sans structure grammaticale, langues à affixes et langues à inflexions (subdivisées en langues synthétiques et analytiques)375.
S’opposant à une typologie stricte à laquelle il préfère un classement graduel376, Humboldt critique l’approche des frères Schlegel, comme il critiquera les typologies bipartite et tripartite. En fait, cette dissension est à replacer au plan de la méthodologie, et, dans le contexte épistémologique du siècle, au plan de la méthodologie biologique : l’approche de Friedrich von Schlegel est une approche anatomique, statique, celui-ci ne prenant en compte que l’organisation interne, la forme générale. Préférant le modèle physiologique à celui de l’anatomie comparée377 utilisé par son compatriote, Humboldt introduit une dimension dynamique dans l’étude de la langue, qu’il envisage comme un organisme vivant et en constante évolution, et non comme un corps mort que l’on dissèque (cf. Jürgen Trabant (1992 : 154)).
Il adopte néanmoins le système schlegelien, tout en y ajoutant une nouvelle catégorie : les langues incorporantes, qu’il supprimera par la suite. Cependant, et malgré les apparences, il s’oppose à la conception organiciste du langage, les objets d’études de la linguistique et des sciences naturelles, étant, selon lui, inassimilables : les langues appartenant au domaine du spirituel et de l’individuel, la généalogie linguistique est un fait historique, et non héréditaire. Dans Ueber die Verschiedenheiten des menschlichen Sprachbaues (1836), il compare les langues, non à des espèces, mais à des entités ou des individualités, et élabore le concept de caractère qu’il oppose à celui de classe. Sa typologie se fonde donc sur les caractéristiques d’individus qui rendent impossible le regroupement en classes. Cependant, dans un souci didactique, il propose le regroupement des langues dotées de propriétés communes en pseudo-classes partielles (cf. Jürgen Trabant (1992 : 161-166)).
Bertil Malmberg (1991 : 270) cite néanmoins l’influence du linguiste allemand Becker – dont la théorie de l’organisme prône une méthodologie de l’unité en opposition au mécanisme, méthodologie de la décomposition378 – sur Humboldt. Adoptant cette thèse que le langage, en tant que phénomène organique, ne peut être apprécié que dans sa globalité – les parties n’étant appréhendables que par leur relation au tout – Humboldt avance que chaque élément n’existe, ne se comprend et ne se définit, que par les autres. Lorsqu’il élabore cette thèse, l’humaniste allemand envisage tout particulièrement le vocabulaire, qu’il considère comme un système structuré, comparant la langue à un tissu379 dont chaque partie est en rapport avec une autre, ou encore, avec la totalité :
‘On peut les comparer [les lois de fonctionnement des langues], en prenant pour modèle les organismes corporels, aux lois physiologiques dont l’observation scientifique requiert un traitement tout autre que celui qui est exigé par la description anatomique des parties prises une à une (Humboldt, Introduction à l’oeuvre sur le Kavi, 24 ; 1835 : 245).’La langue étant envisagée comme un ensemble dont les parties ont une cohérence vivante en étroite liaison avec le tout auquel elles appartiennent ; le mot est par récursivité également appréhendé comme un organisme.
Humboldt fonde sa typologie sur la notion de forme grammaticale (à ne pas confondre avec les formes interne et externe), qu’il considère, dans son point d’aboutissement, comme la fusion de la forme et de la matière380: la définition d’une langue ne doit pas s’appuyer sur un ensemble de caractéristiques, mais sur la combinaison de toutes ses propriétés, qui forment l’unité fondamentale du langage. La flexion des mots, l’isolation, l’agglutination, sont des systèmes d’incorporation des langues, des procédés grammaticaux concernant l’unité du mot et l’unité de la phrase. Cette cohérence organistique est particulièrement prégnante dans les langues flexionnelles, que l’humaniste allemand considère comme une des meilleures expressions, en ce sens qu’elle est la plus aboutie et la plus précise, de la fusion de la forme phonétique et des lois internes de la langue. En effet, la flexion permet, grâce à la construction du mot, de distinguer assignation et dénotation tout en conservant le caractère unitaire du mot.
Dans ses Sprachvergleinchende Untersuchungen (1848)), August Schleicher s’inspire de ce concept humboldtien et pose un double critère de caractérisation des langues : l’expression de la signification et de la relation. La signification est matérialisée par la racine, la modification effectuée sur celle-ci exprimant la relation. Ces composantes et les relations qu’elles inter-entretiennent sont diversement mises en oeuvre381 par les différents idiomes. Adoptant cette perspective comme principe de typologisation des langues, il détermine trois classes, assimilables à celles de ses contemporains : les langues isolantes, agglutinantes et flexionnelles. Tout comme Humboldt, il considère que les langues flexionnelles constituent le point d’aboutissement du système linguistique : réconciliation du matériel et du spirituel, elles possèdent pleinement le caractère molaire caractéristique de tout organisme (cf. Ernst Cassirer (1972 a : 112-115)).
En tant que principe explicatif de la période, le concept d’organisme permet d’articuler les deux phases de ce second moment constitutif de la terminologie. Relevant d’une vision profondément influencée par la pensée idéaliste, il permet d’introduire, par le glissement progressif de sa conception romantique à sa conception « scientifique », une nouvelle positivité dans l’étude des langues. Cette nouvelle positivité est le fruit de la perspective comparativo-historique, dont les frères Schlegel, ou même Humboldt, sont les précurseurs : afin de pouvoir accéder au statut de science, la linguistique rejette la philosophie et adopte le modèle des sciences de la nature.
Autre point d’importance, le concept d’organisme introduit une nouvelle approche du langage : pour établir des parentés entre idiomes, on cesse de se tenir prudemment à la surface des langues, de s’appuyer sur de vagues similitudes apparentes, et c’est par une plongée au coeur de l’anatomie des mots que se révèlent des liens génétiques insoupçonnés. Mais il convient de préciser que l’accession de ce concept au statut de modèle épistémologique est profondément liée à la place centrale qu’il occupe dans la biologie de ce 19e siècle naissant.
La notion d’organisation apparaît dans le dernier quart du 18e siècle. Elle émerge dans la pensée des derniers méthodistes lorsque le botaniste français Antoine Laurent de Jussieu introduit le principe de hiérarchisation des caractères. Pour lui, le caractère, unique ou pluriel, n’est pas un critère fiable. En 1789, dans son Genera plantarum, Jussieu propose alors de remplacer celui-ci par une combinaison hiérarchisée de facteurs. La reproduction étant la fonction essentielle de la fleur, l’embryon (le cotylédon) constituera le critère primaire de classification ; pour cela, il reprend donc la trichotomie établie par John Ray (Methodus plantarum nova (1682)) pour déterminer ses embranchements : les acotylédones, les monocotylédones et les dicotylédones. Le second critère, qu’il emprunte à Tournefort (Éléments de botanique (1694)), est l’enveloppe florale (cf. supra, 2e partie, 9. 2. 3.) ; ce caractère, subordonné au précédent, détermine les sous-embranchements de sa classification (les apétales, les monopétales et les polypétales). Les rapports géographiques entre le pistil (représenté par le formant -gynie) et les étamines sont subordonnées à ce second critère et permettent d’établir les classes. À cette fin, il systématise trois préfixes : épi- (du grec επι - « sur », hypo- (du grec υπο- « sous »), et péri- (du grec περι- « autour de »). Ainsi, les étamines épigynes (stamina epigyna) sont implantées sur le pistil, les étamines périgynes (stamina perigyna) sont implantées autour du pistil, les étamines hypogynes (stamina hypogyna) sont implantées sous le pistil.
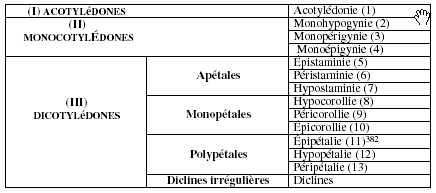
L’embranchement des acotylédones ne comprennent qu’une classe. Celle-ci est donc dénommée en fonction du facteur privilégié – à savoir l’embryon – et se voit attribué le nom d’acotylédonie.
Le second embranchement se subdivise en trois classes. Celles-ci reçoivent donc logiquement le préfixe mono-, rappel mnémonique de la spécificité de l’embranchement (le formant -cotylédonie est écarté pour des raisons mémorielles). Le second facteur est alors utilisé pour différencier les trois classes. Celui-ci leur fournit donc leurs noms : épigynes, périgynes et hypogynes. Le préfixe mono- constitue la marque du premier facteur, constitutif de l’embranchement ; les formants -hypogynie, -périgynie et -épigynie sont la marque du second facteur, constitutif des classes.
L’embranchement des dicotylédones est constitué de quatre sous-embranchements : les apétales (apetalae), les monopétales (monopetalae), les polypétales (polypetalae) et les diclines irrégulières382. Les trois premiers sous-embranchements sont divisés en trois classes : les trois facteurs sont donc nécessaires pour dénommer celles-ci. Comme dans l’embranchement II, la référence à la classe est portée par le second facteur, qui n’apparaît plus que sous la forme du préfixe spatial, différenciateur (comme pour la référence à l’embranchement dans la classe II, le nom n’est pas totalement cité pour des raisons mémorielles). La référence au sous-embranchement se fait par le formant de position 2 : -staminie pour les apétales (elles ne possèdent pas de pétales, l’étamine est donc le facteur de reconnaissance), -corollie pour les monopétales (ce pétale unique est en fait une corolle au pétales indifférenciés, et qui, en tant que pièce unique, n’admet pas le nom de pétale), -pétalie pour les polypétales (où la corolle est composée de pétales différenciés, et autorise donc une telle dénomination). Fidèle à son procédé régressif (mention du nom de l’embranchement dans sa totalité en I, du seul préfixe en II), la référence à l’embranchement est donc in abstentia, par un préfixe 0. D’autre part, la hiérarchie ne nécessite pas de mention explicite à l’embranchement. En effet, la référence au sous-embranchement renvoie implicitement à l’embranchement, puisque seules les dicotylédones possèdent cette subdivision intermédiaire. Le préfixe 0 constitue la marque de l’embranchement ; les formants -staminie, -corollie et -pétalie portent la référence au sous-embranchement ; les formants hypo-, péri- et épi-, qui représentent par rappel mnémonique hypogynie, périgynie et épigynie, sont la marque du troisième facteur, constitutif des classes383.
L’examen des dénominations créées par Jussieu indique que la maîtrise de la hiérarchie classificatoire est nécessaire à l’interprétation des termes. En effet, au fil de la complexification de celle-ci, les facteurs à l’origine des dénominations sont de plus en plus implicites. Ce sont les classes qui permettent de déterminer les sous-embranchements, puis les embranchements. La hiérarchisation des facteurs, matérialisée par la hiérarchisation classificatoire, fait disparaître les mentions des facteurs premiers, puisque que ceux-ci impliquent les facteurs seconds, puis les facteurs de troisième rang384 : la corolle est le taxon constitutif du sous-embranchement des monopétales, sous-embranchement des monocotylédones ; le terme épicorollie renvoie donc automatiquement à l’embranchement des monopétales, et donc, aux dicotylédones. Le principe en germe dans la classification de Linné (cf. supra, 10. 2.) est pleinement exploité par Jussieu.
Pour Vicq d’Azyr, cette hiérarchisation des caractères est entièrement liée à la fonction de ceux-ci au sein de l’organisme. Si le cotylédon est premier dans l’organisation des plantes, c’est en raison de l’importance de la fonction de reproduction dans le monde végétal ; de même, la fonction de nutrition occupe un rôle central dans l’organisation de l’animal (forme des dents, des pieds, de l’estomac, de la langue, des intestins, etc.). Il découle de ce nouveau principe que les caractères de surface sont en étroite corrélation avec ceux de la profondeur de l’organisme (la forme des dents correspond au mode de digestion, etc (cf. supra, 11. 1. 2.)) (cf. Michel Foucault (1966 : 238-245)).
Ainsi, les derniers taxinomistes ne prélèvent plus le facteur classificatoire dans le seul visible, la surface de l’individu étudié, mais dans sa profondeur : en botanique, on troque ainsi les anciens taxons comme la feuille ou la fleur, contre la graine. L’essentialité fonctionnelle, étant devenue un point d’ancrage de la systématique, les taxons n’appartiennent plus au champ du visible, mais à celui de l’organisation interne des individus étudiés385.
La synthèse de toutes ces innovations sera effectuée par Lamarck. Pour lui, la notion d’essentialité fonctionnelle implique que toute modification d’un organe est liée à une réaction de l’organisme aux modifications des conditions de réalisation de ses fonctions. Ainsi, la girafe possède un long cou afin de pouvoir se nourrir. En effet, les organes ayant avant tout une raison d’être fonctionnelle, ceux d’entre eux qui sont inutiles tendent à disparaître, alors que ceux qui sont nécessaires ou essentiels apparaissent ou augmentent de volume. Il n’y a donc pas de classes, mais des successions d’individus qui s’engendrent, et transmettent les modifications adaptatives à la génération suivante.
En raison des liens entre organes, les déviations réactives au milieu peuvent entrer en conflit avec les tendances de renforcement et de complexification propres aux facultés de l’organisme. Celui-ci bloque alors sa transformation. En cas de circonstances favorables (climat, nourriture, dangerosité moindre des prédateurs, présence de refuges, etc.) et si les modifications réactives sont compatibles avec la diversification des facultés de l’organisme, les êtres engendrés sont de plus en plus complexes. La classification animale à partir de critères morphologiques doit être dépassée afin d’établir les liens entre espèces animales, que seule la démarche diachronique permet d’établir (cette démarche est influencée par les études de Lamarck sur les espèces fossiles). L’unité de l’organisme implique l’unité de la vie386 : on passe de la notion d’êtres vivants à celle de vie (cf. Michel Foucault (1966 : 241)).
L’apparition du terme biologie (de βιο - « vie » et -λογια « science ») – créé indépendamment par Lamarck et Treviranus en 1802 – en lieu et place de l’expression physiologie générale (de Φυσιο - « nature » et -λογια « science »), symbolise cette nouvelle approche, qui substitue l’étude de la vie à l’étude de la nature. Sa création simultanée de part et d’autre du Rhin, dans des domaines connexes (Treviranus est naturaliste, Lamarck est zoologiste), révèle l’importance prise par cette conception comme le caractère global de la discipline naissante. Treviranus le définit comme une philosophie du vivant (le terme apparaît dans le titre de son ouvrage Biologie oder die Philosophie der lebenden Natur « philosophie de la nature vivante ») ; dans la pensée de Lamarck, ce terme appartient à la triade météorologie/hydrogéologie/biologie, tripartition de la physique terrestre (science de l’atmosphère, science de l’eau et de la pierre (i. e. de la surface terrestre), science du vivant).
Cependant, si Lamarck fait progresser la discipline, sa notion d’addition d’organes demeure moléculaire, agrégative. Le pas sera franchi par la théorie unitaire de Geoffroy Saint-Hilaire qui fonde son anatomie comparative (Philosophie anatomique (1818), L’Histoire naturelle des mammifère (1819)) sur la loi du balancement des organes (l’hypertrophie de l’un entraîne l’effacement de l’autre), la loi des connexions (les organes conservent toujours entre eux les mêmes relations) et la loi de permanence (aucun organe nouveau ne se crée)387. Toute structure organique n’est autre que l’adaptation au milieu d’une structure commune fondamentale.
Georges Cuvier, dont le fixisme s’oppose au transformisme de Lamarck et de Geoffroy Saint-Hilaire, contribue à la construction d’une conception molaire de l’organisme. Pour cela, il prend en compte les principes directeurs de fonction et de cohérence organique, établissant la dépendance fonctionnelle des organes ; il établit ainsi la loi des corrélations organiques (certains organes excluent ou impliquent la présence d’autres organes)388. En se fondant sur leur organisation interne, il va tenter d’établir les rapports entre les êtres vivants, et publie ainsi en 1817 Le Règne animal distribué d’après son organisation. La fonction ultime des organes est le maintien de la vie, et ils coopèrent afin d’atteindre ce but. Cependant, ils ne possèdent pas tous le même rôle au sein de l’organisme : certains d’entre eux ont une influence prépondérante sur le fonctionnement de l’ensemble (prépondérance que Cuvier accordera au système respiratoire). Ce principe de subordination des caractères, qu’il emprunte à Jussieu, fait de l’organisme, non plus une simple juxtaposition d’éléments, comme dans la théorie de Geoffroy, mais une véritable structure solidaire, dont les parties en interactions forment l’unité.
Si Geoffroy établit l’unité diachronique de l’organisme, Cuvier complétera cette conception en démontrant l’unité synchronique constituée par les éléments de celui-ci : l’anatomie comparée cesse d’être un ensemble de remarques éparses pour devenir la science de l’organisation animale.
La charnière des 18e et 19e siècles sera clairement marquée par le vivant, tant dans les champs scientifique et épistémologique, où il est au centre des préoccupations, que philosophique et linguistique, auxquels il sert de modèle. Lorsque l’on met en parallèle les progrès de la linguistique et de la biologie, on peut constater une communauté de méthodes et de conceptions.
Tout d’abord, on cesse de classifier en ne s’en tenant qu’à la surface des choses. On plonge au coeur du mot comme au coeur de l’organisme. Cette incursion conduit à la prise en compte de la totalité de l’individu étudié : on ne fonde plus sur le caractère unique, ni même sur l’ensemble des caractères, mais sur la combinaison des propriétés de l’objet (Jussieu, Humboldt). Cette combinaison se traduit par les interactions entre les parties de l’objet d’étude, qui cesse d’être une somme pour devenir une symbiose. La modification de l’une de ses parties entraîne celle d’une autre (loi du balancement des organes de Geoffroy, interaction entre forme externe et forme interne de Humboldt). Cette synergie se traduit également par la hiérarchisation des éléments (rection de la racine par la flexion, loi de subordination des caractères de Cuvier).
Émerge donc une conception molaire de l’individu linguistique et biologique, qui, par une analyse récursive, se voit appliquée à l’ensemble auquel appartient cet individu. Celui-ci est mis en perspective : perspective génétique (typologie historique de August-Wilhelm Schlegel, conception transformiste de Lamarck et Geoffroy Saint-Hilaire) ; perspective réticulaire (liens de parenté de Lamarck, conception tissulaire du lexique par Humboldt).
L’objet d’étude n’est donc plus immanent, mais le fruit d’une construction réactive. L’espace intérieur et extérieur ne sont plus cloisonnés, mais en interactions constantes : l’expérience sensorielle est retraitée par le langage, comme elle est traitée par l’organisme qui se modifie afin de s’adapter au milieu. Organisme et langage sont en constante modification pour adapter les organes à leur fonction (la notion de réactivité de Lamarck), le son à la pensée (l’Energéia de Humboldt). C’est par cet incessant travail d’adéquation du visible à l’invisible que l’organe de surface/la forme externe permet de deviner l’organe/la forme interne. Comme le langage prépare et raffine l’expérience sensorielle, l’organisme fait en sorte que celle-ci ne soit pas une agression pour le corps.
La notion d’interaction sera donc la grande conquête épistémologique de cette période charnière entre l’âge classique et le monde moderne : l’interaction interne de la synthèse et l’interaction externe de la réticulation.