1.1.3. Rôle de la voie Magnocellulaire et Parvocellulaire dans l’intégration d’une forme
Les axones des cellules ganglionnaires convergent à la surface de la rétine pour former le nerf optique qui se projette principalement sur les corps genouillés latéraux dont la structure est organisée en six couches. L’organisation des champs récepteurs du CGL est identique à celle des cellules ganglionnaires. Mais, la taille des champs récepteurs, à partir de la rétine, augmente au fur et à mesure des différentes étapes de traitement (CGL, V1, V2, etc…).
Les cellules en parasol se projettent dans les couches magnocellulaires (les deux couches inférieures), les cellules naines dans les couches parvocellulaires (les quatre couches supérieures), et les cellules bi-stratifiées dans les inter-couches koniocellulaires. Ces destinations anatomiques ont donné leur nom à trois voies visuelles : la voie magnocellulaire, la voie parvocellulaire et la voie koniocellulaire (Lee, 1996). Nous nous intéresserons plus particulièrement aux voies magnocellulaire et parvocellulaire car elles sont dépendantes du contraste de luminance. Les cellules bi-stratifiées, qui forment le trajet koniocellulaire, ont des champs récepteurs de grande taille, et répondent aux fréquences spatiales basses. Mais, la voie K n’est pas sensible aux stimulations achromatiques (Hendry & Reid, 2000 ; Kaplan, 2003). C’est pour cette raison que cette voie ne sera pas développée ici.
Tout signal visuel est donc précocement analysé par ces deux systèmes qui ont des propriétés distinctes et complémentaires. Le système magnocellulaire (ou voie M) répond à des hautes fréquences temporelles (image en déplacement rapide sur la rétine ou papillotement rapide de tâche lumineuse) et aux basses fréquences spatiales 2 (l’exemple d’une image filtrée est donnée Figure 4). Ce dernier répond très rapidement, 50 ms après l’apparition et à l’extinction d’un signal visuel. En d’autres termes, cette voie permet de signaler efficacement les changements brusques et rapides de luminance. Le système parvocellulaire (ou voie P) possède au contraire une résolution temporelle basse et répond aux fréquences spatiales élevées. Les cellules de cette voie répondent préférentiellement à un contraste stable dans leur champ récepteur. Cette voie est activée plus tardivement (70 ms après l’apparition du signal), et véhicule des informations chromatiques et des détails de l’image, son contour par exemple (Kaplan, 2003).

Les différences de sensibilité spatiale et temporelle, des voies M et P, ont été démontrées dans une série d’études où des inhibitions spécifiques aux deux voies étaient réalisées. Merigan et al. (1990, 1991) ont réalisé une lésion partielle dans la voie M ou P du CGL de deux singes, et une étude comportementale a ensuite été effectuée. Merigan et Maunsell (1990) ont inhibé spécifiquement la voie M. Dans ce cas, les deux animaux ne répondaient pas à des fréquences temporelles élevées (10 Hz) lorsque ces dernières étaient présentées dans le champ visuel affecté par la lésion. Par ailleurs, des réponses correctes étaient enregistrées lorsqu’un réseau de 2 cdp était présenté (correspondant à une fréquence spatiale élevée). Merigan, Katz et Maunsell (1991) ont par la suite montré qu’une lésion de la voie P réduisait considérablement les réponses comportementales pour un réseau de 2 cpd. De même, les réponses n’étaient pas affectées par la présentation de fréquences temporelles élevées (10 Hz). Allison, Melzer, Ding, Bonds et Casagrande (1999) ont montré que les réponses des cellules simples de V1 aux contrastes étaient affectées par une inactivation de la voie M ou P. Une inhibition de la voie M dans le CGL réduisait la réponse des neurones pour l’ensemble des niveaux de contrastes. Par contre, l’inhibition de la voie P n’avait aucun effet sur des réseaux dont le contraste était inférieur à 20%. Ces deux voies agissent de façon parallèle mais elles sont confondues à la sortie de V1.
Les premières études physiologiques ont montré que les cellules de la voie M étaient beaucoup moins représentées autour de la fovéa, comparativement aux cellules de la voie P. Toutefois, des études plus récentes ont montré que le ratio des cellules naines et en parasol était identique dans la rétine du singe (Perry, Oehler & Cowey, 1984 ; Silveira & Perry, 1991). Ces résultats mettent ainsi en doute la sélectivité des voies M et P pour des stimulations où l’excentricité du signal varie. Toutefois, une étude de Dacey et Petersen (1992), chez l’observateur humain, n’a pas corroboré ces résultats. En effet, ils ont observé que le ratio des voies P/M dans la rétine était de 30/1.
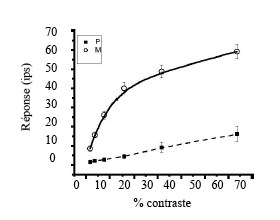
Une distinction supplémentaire entre les deux systèmes concerne la taille des champs récepteurs 3 . En effet, le diamètre des champs récepteurs augmente entre la fovéa et la périphérie pour les deux voies, mais quelle que soit l’excentricité, le diamètre des champs récepteurs des cellules magnocellulaires est toujours plus large que celui des champs récepteurs des cellules parvocellulaires (Croner & Kaplan, 1995). Cette différence de taille entre les deux voies a une conséquence importante, puisque la sensibilité au contraste des deux systèmes n’est pas la même. Ainsi, lorsqu’on enregistre la réponse des deux types de cellules pour des contrastes de luminance qui augmentent pas à pas, les résultats montrent une réponse élevée des cellules magnocellulaires pour des contrastes faibles et une réponse faible des cellules parvocellulaires pour tous les niveaux de contraste (Kaplan & Shapley, 1986). Par ailleurs, le gain de contraste 4 des cellules de la voie M augmente rapidement pour les premiers niveaux de contraste, les cellules sont ensuite saturées pour un contraste excédant 15-20 %. La réponse des cellules de la voie P, au contraire, augmente de façon graduelle. Il est tout à fait possible que les cellules de la voie M soit déterminées par deux mécanismes : un pour les plus petits niveaux de contraste et l’autre pour des niveaux de contraste élevés (Kaplan, 2003). La Figure 5 montre la réponse des deux populations de neurones selon le niveau de contraste. De la même façon que pour les canaux M et P, Lee, Pokorny, Smith, Martin et Valberg (1990) avaient observé des réponses aux contrastes différentes pour les cellules ganglionnaires en parasol et naines de la rétine.