2.2.3.2. Quelle est la réponse des neurones du cortex visuel à une surface uniforme ?
Les expériences précédentes ont permis de montrer des activations des neurones de V1 lorsqu’un signal est présenté dans une région du champ visuel recouvrant la tache aveugle (Fiorani et al., 1992) ou les scotomes (Pettet & Gilbert, 1992). Ces résultats permettent de conforter l’hypothèse de Spillman et Werner (1996) selon laquelle les connexions horizontales seraient fortement impliquées dans le processus de remplissage. Au debut de ce chapitre, nous avons vu que de nombreux auteurs (Durgin, Tripathy & Levi, 1995 ; Gerrits & Vendrik, 1970 ; Grossberg & Mingolla, 1985 ; Walls, 1954) émettaient l’hypothèse selon laquelle ce processus expliquerait le codage de la surface. L’objectif est ici de déterminer si le remplissage participe directement au traitement de la surface.
La luminosité 11 d’une surface peut facilement être manipulée par rapport au niveau réel de luminance par une technique d’induction. Par exemple, lorsqu’un carré gris moyen est entouré d’un fond noir, le carré paraît plus clair qu’il ne l’est quand il est perçu de façon isolée, et inversement lorsqu’un fond plus clair est présenté. Un exemple est montré à la Figure 16.

Rossi, Rittenhouse et Paradiso (1996) ont utilisé cette technique d’induction mais avec des stimuli différents. Dans leur condition « induction », une surface centrale était entourée par deux autres régions (à droite et à gauche) dont la luminance variait dans le temps du clair au sombre. Dans une autre condition, les deux surfaces étaient statiques avec un niveau gris moyen, mais alors, c’est la luminance de l’aire centrale qui variait du clair au sombre. Une condition contrôle était utilisée dans laquelle aucun autre stimulus n’était présenté, mais la luminance des deux surfaces adjacentes variait du noir jusqu’à un niveau de gris moyen. Les auteurs enregistraient chez le chat les neurones de l’aire visuelle primaire qui avaient leurs champs récepteurs situés dans la surface centrale. Les résultats ont indiqué que dans la condition d’induction, 75 % des neurones (120 sur 160) étaient activés par les variations de luminance du pourtour, et ce malgré le fait que les variations de luminance se produisaient en dehors du champ récepteur enregistré. La variation de luminance de la région centrale produisait aussi une réponse, mais qui était plus importante lorsque la luminance était claire. Dans la condition où l’aire centrale était absente, la décharge des neurones était équivalente à une décharge spontanée. Cette recherche montre un résultat très important pour la question du traitement de la surface, à savoir que la luminosité est traitée à partir de V1. Mais ici, les neurones n’agissent pas forcément comme des détecteurs de luminance puisque le pourtour peut être la dimension qui contribue à leur activité. Un résultat identique a été montré lorsque le champ récepteur des neurones enregistrés était situé dans un Mondrian 12 (MacEvoy & Paradiso, 2001). Ce résultat a été répliqué, par Kinoshita et Komatsu (2001), chez le singe avec une procédure simplifiée : la luminance du fond était fixe, et, seule la luminance du carré variait.
Rossi et Paradiso (1999) ont voulu déterminer si les réponses neuronales observées dans l’étude précédente, se produisaient uniquement dans le cortex ou si des réponses similaires étaient observées à une étape plus précoce du traitement visuel. Avec les mêmes stimuli que dans l’étude de Rossi et al. (1996), ils ont examiné la réponse de 75 neurones du CGL. Lorsque la luminance variait en dehors de l’aire centrale, la réponse des neurones n’était pas aussi élevée que dans le cortex, puisque 42 neurones (soit seulement 56 %) produisaient une activation significative. Par ailleurs, ils ont utilisé la même procédure expérimentale, mais cette fois pour enregistrer 33 cellules ganglionnaires dans la rétine grâce à un traceur optique. Mais, dans ce cas, aucune activité n’était constatée. Cette étude suggère donc que l’information de surface est intégrée à travers le champ visuel au niveau du cortex, voire dès le CGL, puisque la perception de la luminance est influencée par les variations de luminance des surfaces voisines, même éloignées.
Notre question est de déterminer l’implication du processus de remplissage dans le traitement de la surface. Pour cela, la taille du signal était manipulée. Selon le raisonnement suivant : la réponse des neurones dont le champ récepteur est situé au centre de la figure, devrait diminuer avec l’augmentation de la taille de la stimulation. En effet, augmenter la taille du signal revient à ralentir la propagation de l’information le long des connexions horizontales puisque la distance à parcourir augmente. Komatsu, Murakami et Kinoshita (1996) ont enregistré chez le macaque 29 neurones situés dans l’aire V1 pendant que plusieurs carrés de tailles différentes (plus petits ou plus larges que la taille des champs récepteurs, de 0.3 à 6.7 deg) étaient présentés. Les résultats ont montré un déclin de la réponse lorsque les stimuli devenaient de plus en plus larges. En d’autres termes, plus le contour s’éloigne, moins il y a de remplissage, sauf pour 1/3 des neurones pour lesquels l’activité restait constante quelle que soit la taille (ces neurones étaient situés à des profondeurs différentes dans le cortex). Par la suite, MacEvoy, Kim et Paradiso (1998) ont voulu déterminer la taille critique au-delà de laquelle l’information de surface n’influencerait plus les réponses des cellules dont le champ récepteur est situé à l’intérieur de la surface. Pour cela, ils ont utilisé un disque dont la taille augmentait, et le champ récepteur était positionné au centre. Les résultats ont indiqué qu’une majorité des neurones (72 %) étaient activés par la présentation d’un disque lumineux, même quand les contours de ce disque étaient placés en dehors du champ récepteur de la cellule. L’augmentation de la taille du disque avait pour effet principal de diminuer la réponse des neurones jusqu’à une taille critique (8 deg) pour laquelle les cellules n’étaient plus activées. Par ailleurs, les auteurs ont également constaté d’autres types de réponses neuronales. Dans certains cas, l’activité neuronale diminuait avec l’augmentation de la taille du disque, mais contrairement au résultat précédent, ces cellules restaient toujours actives. Un troisième résultat indiquait que l’activité des neurones augmentait avec la taille du disque, pour des tailles allant jusqu’à 30 deg. Par la suite, MacEvoy et al. (1998) ont remplacé le disque par un réseau sinusoïdal, dont l’utilisation est plus conventionnelle. Les résultats ont indiqué que seule la moitié des neurones étaient affectés par la variation de taille, mais pour ces neurones, les résultats obtenus étaient similaires à la condition où un disque uniforme était utilisé. L’hétérogénéité de ces résultats semble indiquer que chaque cellule n’intègre pas simultanément la surface. L’hypothèse du remplissage est ici controversée puisque malgré les grandes tailles du signal, certains neurones sont toujours activés (Komatsu, Murakami & Kinoshita, 1996 ; MacEvoy, Kim & Paradiso, 1998). Ces résultats montrent que le phénomène de remplissage n’est pas suffisant à lui seul pour rendre compte de la perception d’une surface. Ces résultats sont généralisables à l’observateur humain, car la perception de la luminance est similaire à celle du macaque (e.g. Huang, MacEvoy & Paradiso, 2002).
Ces études (Rossi & Paradiso, 1999 ; Rossi, Rittenhouse & Paradiso, 1996) sont proches des effets contextuels, déjà cités dans le premier chapitre à la différence près que la cible et le contexte sont non plus des barres mais des surfaces uniformes. L’utilisation des effets contextuels est en effet la seule méthode capable de rendre compte du traitement de la surface, puisqu’en effet, une surface n’est jamais perçue de façon isolée. Ces expériences ont montré que les cellules de V1 sont activées par une surface uniforme.
Pour tester l’hypothèse du rôle du remplissage dans le codage de la surface, de nombreux auteurs ont manipulé la taille du signal (Komatsu, Murakami & Kinoshita, 1996 ; MacEvoy, Kim & Paradiso, 1998). Ainsi, lorsque l’on enregistre des cellules dont le champ récepteur est positionné au centre de la figure, et si la taille du signal augmente alors on pourrait s’attendre à ce que la réponse de ces cellules diminue. Toutefois, ce type de réponse n’est pas observé pour l’ensemble des neurones, en effet certaines cellules maintiennent une activité constante. Par conséquent, le processus de remplissage dans le traitement de la surface semble être fortement mis en jeu, mais les variations d’activité des cellules ne permettent pas d’affirmer que ce processus définit totalement le traitement de la surface (Gilbert, 1996).
Les expériences qui ont été développées dans les paragraphes précédents (Rossi & Paradiso, 1999 ; Rossi, Rittenhouse & Paradiso, 1996) repose sur un contraste entre deux surfaces. Ces études ne déterminent pas l’influence des contours dans la réponse de V1 à la luminosité d’une surface. Hung, Ramsden, Chen et Roe (2001) ont étudié l’implication des contours dans la perception de luminance. Les auteurs se sont intéressés à l’activité de la surface dans le cas de l’effet Craik-O’Brien-Cornsweet (appelé également effet COCE). Cet effet est considéré comme une illusion où la luminosité d’une région est affectée par la discontinuité de luminance au bord de cette région (un exemple est donné à la Figure 17). Dans ce cas, il n’y a pas de contraste entre les surfaces, car la différence de luminosité est interprétée à partir du changement produit par les bords. Hung et al. (2001) ont également utilisé deux autres types de stimuli : deux surfaces contiguës dont la luminance changeait réellement, et deux surfaces qui étaient équiluminantes, avec un profil de luminance aux bords qui augmentait ou diminuait (« Narrow Real Stimulus »). Ce dernier stimulus se distingue du COCE par le fait que les variations de luminance aux bords des deux surfaces ne se font pas graduellement, ce qui a pour effet d’inverser la luminosité par rapport au COCE. Les profils de luminance et de luminosité des stimuli sont présentés à la Figure 17. L’utilisation de ces trois types de stimuli permettrait aux auteurs de déterminer la contribution des contours dans la perception de luminance.
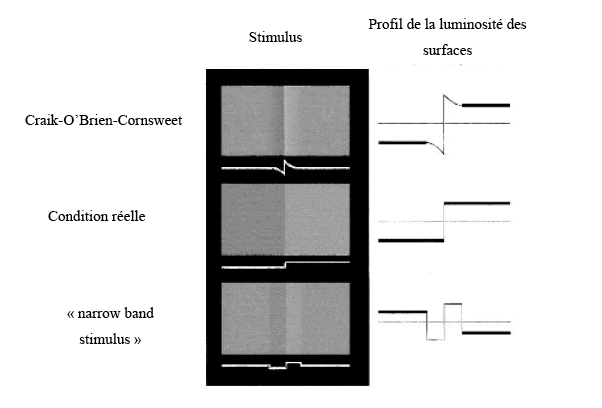
Hung et al. (2001) ont utilisé la technique d’image optique, mais ils ont également effectué un enregistrement de la réponse des neurones dans les aires V1 et V2 du chat. Pour s’assurer que les neurones ne répondaient pas à un contraste, le contour, qui sépare les deux surfaces, était placé à une distance minimale de 1 deg du champ récepteur. Les résultats obtenus avec la méthode de l’image optique, ont indiqué une activité plus importante dans V2 que dans V1 pour les trois types de stimuli. De plus, une forte activité était notée pour les stimuli dont la luminance changeait réellement. Par contre, l’activité neuronale était plus faible quelle que soit l’aire corticale pour l’effet COCE et le « Narrow Real Stimulus ». Toutefois, ces résultats sont à relativiser puisque les enregistrements ont été réalisés chez le chat et non chez le primate. En effet, il existe des différences fondamentales entre le primate et le chat, notamment il existe des arguments pour supposer qu’il n’y a pas de voie parvocellulaire chez le chat (Rodieck, 1998).
Les études précédentes (Rossi & Paradiso, 1999 ; Rossi, Rittenhouse & Paradiso, 1996) ont montré que les neurones du cortex visuel primaire répondaient à un contraste de luminance entre deux surfaces. Mais, l’origine de cette réponse est incertaine : est ce que les neurones sont activés via les bords de la stimulation ou bien directement à partir des caractéristiques de la surface ? Cette réponse n’a pas trouvé de réponse totalement satisfaisante. Par contre, dans l’effet COCE, le contraste de surface est absent et le changement de luminosité est le résultat d’un contraste. Les résultats de Hung et al . (2001) semblent être plus robustes dans V2 que V1. Ainsi, il faudrait attendre V2 pour que la luminosité d’une surface soit induite depuis un contour. De plus, les auteurs suggèrent que la perception de la luminosité est encodée par l’aire V1 pour un contraste entre deux surface, et par l’aire V2 pour un contraste induit à partir des bords. En d’autres termes, cette partie nous a permis de montrer qu’il existait des cellules qui sont sensibles à la luminance d’une surface mais cette réponse peut être accentuée à partir du moment où il y a une induction de la luminosité depuis les contours.