Les « interprétations chimiques »
Soit donc les interprétations chimiques de la croissance. Teissier rappelle d’abord qu’en 1906, le biologiste et embryologiste germano-américain Jacques Loeb (1859-1924) avait admis le fait que la synthèse des nucléines dans l’œuf en segmentation était soumise à deux influences contradictoires : « l’une est la loi d’action de masse qui veut, puisque l’œuf ne reçoit aucune nourriture du milieu extérieur, que la vitesse de formation des noyaux diminue à mesure que leur nombre augmente ; l’autre est la présence d’une catalyse nucléaire qui fait que la vitesse augmente proportionnellement au nombre de noyaux déjà formés. » 120 On obtient donc l’équation différentielle « logistique » 121 suivante dont la formulation, dans ce même contexte biologique, est proposée par le physiologiste T. Brailsford Robertson en 1908 122 :
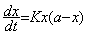
Ainsi, Robertson redécouvre de son côté la courbe en S. Il l’appelle la courbe « autocatalytique » en référence aux réactions chimiques du même nom. Mais, selon lui, le fait qu’elle décrive aussi bien la croissance de noyaux cellulaires que celle des plantes, des animaux et de l’homme, indique qu’il existe chez les êtres vivants une substance biochimique autocatalytique unique, à l’origine de tous les processus de croissance 123 . Selon Teissier, en revanche, le phénomène biologique est certes « ramené » par là à « un phénomène bien connu dans la chimie des ferments » 124 . Mais alors que cette assimilation de la croissance biologique à un phénomène chimique était « légitime » 125 pour le cas des noyaux cellulaires et des cytoplasmes, Teissier conteste vigoureusement son emploi pour des organismes entiers 126 . Si, au niveau cellulaire, la loi mathématique de Loeb est valable biologiquement, c’est parce que son interprétation chimique est biologiquement fondée et fonde en retour la traduction mathématique. La loi mathématique est donc biologiquement valable et elle abandonne son simple statut de « description quantitative » en devenant une « loi de la croissance » authentique à partir du moment où elle est légitimement interprétée et biologiquement construite. Cependant, Teissier rappelle que de nombreux biologistes ont, sans égard pour cette condition, ajusté empiriquement cette loi quantitative pour traduire des croissances de l’organisme entier. Il se dit rigoureusement opposé à ce genre de procédé. L’argument de Teissier en défaveur de l’emploi de cette loi pour l’organisme tient au fait que l’hypothèse d’un système clos ne tient plus du tout dans le cas d’un animal qui trouve de quoi se nourrir dans son milieu extérieur, cela tout au contraire de l’œuf 127 . Comme l’interprétation chimique ne vaut plus à cette échelle, ce n’est donc plus une loi quantitative interprétable pour lui. Donc « on tombe dans le pur arbitraire » 128 , selon Teissier, si on s’ingénie à l’employer pour l’organisme entier.
Mais Teissier se fait tout de même une objection : pourquoi cette loi, qui n’est rationnellement légitime qu’à l’échelle cellulaire, vaut-elle encore empiriquement à l’échelle de l’organisme, ou des organes, lorsqu’on la complexifie un peu ? Parce que, selon lui, Robertson retrouve ce faisant, mais sans s’en apercevoir, la vieille loi logistique de Pierre-François Verhulst (1804-1849) applicable à la croissance des populations et remontant à 1838. Or, dès 1925, le physicien et chimiste Alfred J. Lotka (1880-1949) a montré qu’elle « représentait le mode d’accroissement le plus simple d’un agrégat cellulaire » 129 . En ce sens, Teissier rappelle qu’il existe un pont indéniable entre les lois de la croissance des agrégats de cellules, à « ontologie métrique » pourrions-nous dire, et les lois de la multiplication, à « ontologie populationnelle », de la dynamique des populations 130 . Teissier souligne que cette possible identification (valable pour les cas de croissance d’agrégats très simples) a d’abord été inaperçue, notamment par Robertson. Elle permet pourtant de légitimer en partie le recours à la loi de Loeb pour l’organisme entier.
C’est alors l’occasion pour Teissier d’énoncer une remarque épistémologique d’importance : « une formule a toujours un contenu beaucoup plus riche que ne le croit celui qui l’a établie » 131 . Autrement dit, la formulation mathématique aurait toujours la faculté de masquer les sources de sa véritable « légitimité » aux yeux mêmes de celui qui la produit. Comme dans un authentique mouvement dialectique, la première donation de signification en date ne s’identifierait pas avec la signification effective de la formule, c’est-à-dire avec son contenu biologiquement significatif. Cette signification effective de la « formule » mathématique ne se révèlerait qu’après une nécessaire critique des premiers travaux de description et de formulation. Selon Teissier, cette critique doit s’armer du critère de la signification biologique et physiologique des éléments qui interviennent dans la « formule ».
Mais pour autant, et cela est important à noter, à la différence du cas traité par Loeb, Teissier ne considère pas que cette similitude puisse être tenue pour une interprétation totalement légitime. Rapporter la croissance d’un organe ou d’un organisme à la dynamique des populations cellulaires dont il est composé ne constitue pour lui en rien une légitimation définitive. Car il n’y a là qu’une similitude « formelle » qu’il juge « artificielle » 132 :
‘« C’est à ses qualités formelles, qui n’ont strictement rien à voir avec l’autocatalyse, que la formule de Robertson doit sa valeur pratique … » 133 ’C’est bien pour Teissier une critique d’« ordre biologique » : ceux qui croient avoir interprété chimiquement la loi logistique pour l’organe ou l’organisme se rendent coupables de supposer que l’être vivant se réduit à quelque chose comme une « substance vivante » assimilable à une « masse amorphe et homogène » 134 et de composition chimique très simple. Le succès relatif de ces lois de croissance pour l’organisme entier ne tient pas au fait qu’elles sont interprétables chimiquement, comme veulent nous le faire croire des chercheurs comme Robertson, mais seulement au fait qu’elles sont formellement identiques à des lois connues mais très formelles (car reposant sur une homogénéisation complète des individus d’une population) de la dynamique des populations.
Pour Teissier donc, en biologie, l’identité formelle ne légitime pas en soi le recours à une loi quantitative au niveau théorique quand bien même, au niveau pratique, elle peut avoir un intérêt. Ainsi, pour lui, alors que l’interprétation populationnelle des lois de croissance chez les animaux supérieurs est une légitimation très imparfaite parce que formelle, les théories chimiques de cette même croissance biologique macroscopique ne donnent en revanche rien d’autre que « des schémas à peu près vides de tout contenu concret » 135 . Il met cette complète erreur sur le compte d’un « mécanisme outrancier », et il assimile le travail que font ces théories chimiques de la croissance globale à l’étude d’une pure « représentation verbale dont la rigueur apparente ne résiste pas à la plus simple critique » 136 . Teissier vise donc ici au premier chef le « mécanisme » qui semble manifestement revendiqué par ailleurs par Jacques Loeb dans son ouvrage populaire The Mechanistic Conception of Life : Biological Essays, paru en 1912 137 . Dépourvues de toute interprétation, même au niveau formel, ces « formules » chimico-mathématiques ne sont pour Teissier que « métaphores brillantes » 138 , certes parfois utiles, mais stérilisantes à terme pour la pensée biologique. On notera ici la fréquence des termes péjoratifs renvoyant au caractère purement verbal, voire linguistique (comme le mot « métaphore »), d’une représentation qui tourne à vide, d’un jeu sur les mots qui n’a donc pas de prise concrète sur la réalité biologique. Il tire de cette mise en garde générale une condamnation des approches d’emblée physicochimiques de la croissance organique. Même si l’objectif réductionniste peut en être, à terme, légitime dans le cadre d’un matérialisme conséquent qu’il prône par ailleurs 139 , cette interprétation risque d’être très longtemps encore prématurée 140 . Selon lui, il ne faut pas croire que l’on puisse dès aujourd’hui la formuler.