Les « lois élémentaires » de la croissance
Pour Teissier, il faut donc commencer par l’expression physiologiquement raisonnée de lois quantitatives élémentaires. C’est Jacques Monod (1910-1976) 155 , alors inscrit en thèse à la Sorbonne, qui a mené ces recherches avec Teissier en 1935. Monod et Teissier, en cultivant des microorganismes appelés Glaucoma piriformis et en prenant en compte la variation de la relation entre le taux de division de ces microorganismes et la concentration en substances nutritives du milieu de culture, avaient déjà obtenu une formulation de la loi logistique plus complexe et permettant de traduire une des premières particularités physiologiques des êtres pluricellulaires 156 .
Mais Teissier considère que l’on doit aussi prendre en compte « le fait que les déchets qui s’accumulent dans le milieu de culture exercent une action inhibitrice sur la multiplication des cellules » 157 . On ne peut pas non plus négliger le fait que ces microorganismes, comme les animaux supérieurs, développent une « énergie d’entretien » et une « énergie de croissance » 158 . En conséquence, afin de tenir compte de ces autres relations physiologiques élémentaires entre les cellules tout en ne complexifiant pas trop d’emblée l’objet d’étude, il est préférable de s’attaquer ensuite à la croissance des cultures de tissus organiques. Les tissus peuvent en effet être considérés aussi bien comme des « organismes rudimentaires que comme des populations cellulaires » 159 . Ils permettent donc d’insérer délicatement et progressivement du physiologique dans du populationnel. Selon la perspective de Teissier, les lois quantitatives de la dynamique des populations, d’abord trop formelles, vont ainsi gagner en réalisme physiologique grâce à cette recherche empirique qui se place délibérément aux frontières des objets d’étude et des disciplines.
Teissier évoque ensuite son travail avec Boris Ephrussi (né en 1901) effectué à l’université Caltech en 1930 et publié en 1931 160 : l’étude de la croissance résiduelle 161 des cultures de fibroblastes 162 . Cette croissance ne se produit qu’aux dépens des réserves accumulées par les cellules. Tout en manifestant un phénomène véritablement physiologique, elle est donc plus simple à formuler car on peut supposer qu’elle ne dépend que de deux facteurs : la quantité de réserves disponibles et le nombre de cellules susceptibles de se diviser. Ephrussi et Teissier arrivent à la « traduction » de cette croissance sous forme d’une équation différentielle simple, soluble et dont l’ajustement aux observations est très bon. Une telle introduction des paramètres physiologiques peut ensuite s’étendre aux plantules 163 essentiellement parce qu’on peut aussi les faire croître en ne leur conférant que des ressources limitées. Teissier en tire une conclusion importante : « Dans le cas des plantules, comme dans celui de la croissance résiduelle, nous avons pu pousser notre analyse jusqu’à son terme logique : l’énoncé d’une loi quantitative aussi facile à traduire en langage biologique qu’en langage mathématique. » 164 Ainsi, pour Teissier, on a dans ce cas réellement affaire à une loi quantitative de la croissance physiologiquement (donc légitimement) interprétée parce que tous les paramètres introduits ont « une signification biologique précise » 165 . C’est la simplicité du matériel étudié qui, selon Teissier, a permis cette traduction quantitative légitime 166 . Cette loi peut s’étendre, si on la complexifie encore, à la croissance de l’embryon de truite. La complexification physiologique et mathématique consiste dans ce cas à considérer que les réserves du sac vitellin ne servent pas seulement à la croissance mais aussi partiellement à l’entretien de l’embryon. Pour le poids x de l’embryon et la quantité y de réserves, on obtient alors les équations suivantes :
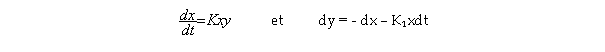
Ces équations ne conduisent malheureusement pas à une solution analytique simple 167 . Et Teissier déplore qu’on ne puisse l’exprimer à l’aide de fonctions élémentaires. Selon lui, l’attaque par le calcul numérique approché donne cependant une courbe de croissance conforme aux faits observés.
Ce qui est au fond également instructif dans ce cheminement que nous communique Teissier, c’est qu’il y est souligné l’idée qu’on ne peut traduire mathématiquement des lois physiologiques de la croissance que si l’on intègre dans cette traduction mathématique une complexité analytique de même ordre, ou en tout cas croissant dans le même sens, que celle du substrat biologique. Et chaque paramètre nouveau ou chaque relation nouvelle entre les variables doit rendre compte d’un fait physiologique précis et auparavant conceptualisé et verbalisé dans le langage de la physiologie et de la biologie. Si l’on voulait passer à l’embryon du mammifère, comme celui de la souris par exemple, Teissier indique qu’il faudrait prendre en compte un nouveau fait physiologique : les substances nutritives viennent dans ce cas soit de la mère, au début de la croissance, soit du milieu extérieur, par la suite. Or, il existe des mesures expérimentales sur la souris. Teissier précise qu’il a reculé devant la difficulté mais sans en expliquer outre mesure les raisons : « on rencontrerait sans doute dans cette étude de très graves obstacles, dont il est d’ailleurs bien inutile de vouloir imaginer la nature » 168 . Nous pouvons conjecturer de notre lecture des pages précédentes qu’ils seraient de nature mathématique. Non seulement les équations différentielles ne donneraient pas lieu à des solutions analytiques, mais sans doute n’y aurait-il même pas moyen d’écrire des équations différentielles, c’est-à-dire de proposer une traduction mathématique analytique des relations physiologiques impliquées.
La recherche de lois quantitatives pour la croissance peut donc sembler être dans une impasse sévère à cause de ce mur de la complexification mathématique des traductions des faits physiologiques. À la fin de son premier chapitre, Teissier dramatise un peu le tableau de cette situation pour nous préparer à saisir ce qu’il va nous présenter, ensuite et pour finir, comme une issue partielle mais déjà victorieuse. C’est ce moment qu’il choisit pour nous présenter (toujours dans une chronologie rationalisée a posteriori) une issue inattendue, pourrait-on dire, celle de la « croissance relative ». Ce règlement du problème de la complexification des traductions, un peu biaisé du fait qu’il contourne curieusement l’obstacle au lieu de l’affronter, mérite d’être restitué dans ses choix techniques et épistémologiques car, dans la biologie de la morphogenèse et, en particulier, dans la biologie française, il constituera une sorte de paradigme de ce que l’on appellera plus tard un « modèle mathématique ».