Des modèles de la génétique au « module » des chromosomes
La question initiale qui oriente le travail de DEA est celle de savoir s’il est possible de mieux caractériser le caryotype, ou profil chromosomique, de certaines graminées : les ray-grass anglais ou Lolium perenne. Si l’on veut pouvoir produire des hybrides raisonnés et eux-mêmes fertiles de toutes plantes, il est en effet indispensable de connaître et de savoir reconnaître précisément les homologies de structure entre les chromosomes des deux génomes parentaux 1400 . À défaut de telles homologies, l’appariement des chromosomes parentaux ne pourra s’effectuer correctement. Or, en ce début des années 1970, les cytogénéticiens sont face à des données très variables et peu sûres. Au microscope optique, et même avec les techniques de traitement et de coloration, la morphologie des chromosomes apparaît très variable et n’est que très schématiquement représentable 1401 . On sait que la variabilité des longueurs absolues apparentes des chromosomes tient à leur propriété d’être spiralés, conformément à une idée déjà proposée par le botaniste allemand Baranetzki en 1880 et précisée entre-temps par le biologiste britannique Cyril Dean Darlington (1903-1981) en 1937 1402 . Mais la seule parade proposée consiste à établir le tableau des différentes longueurs caractéristiques du caryotype, ou caryogramme, en longueurs relatives. Ainsi, pour chaque cellule, on détermine d’une part les rapports de longueurs entre les bras de chacun de ses chromosomes (rapports centrométriques), d’autre part les rapports entre les longueurs de ses chromosomes (longueurs relatives).
C’est là que la suggestion propre à de Reffye et Essad intervient. Lorsqu’elle est publiée, de Reffye est déjà en poste en Côte-d’Ivoire depuis deux ans. Cette contribution, même modeste, semble tout de même significative car elle fait intervenir la notion de « module », tout en prêtant aux mathématiques élémentaires qui l’accompagnent un rôle descriptif sans qu’elles soient pour autant statistiques. Elle ne sera pas négligeable pour la conception que de Reffye se fera de la modélisation des formes dans les sciences du vivant. Qu’en est-il exactement ?
Philippe de Reffye et Sadi Essad proposent qu’à l’aide de la formule mathématique classique
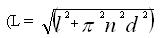
exprimant la longueur développée L d’une spirale à partir de sa longueur apparente l, du nombre n et du diamètre d de ses spires 1403 , on s’attache à chercher les longueurs relatives développées, c’est-à-dire réelles, des chromosomes. Dès lors que l’on a repéré le nombre de spires de la spirale, il est en effet possible de faire le calcul. Les longueurs ainsi développées s’avèrent remarquablement stables : les coefficients de variation passent de 30% à 5% et les écart-types obtenus sont dans la marge d’erreur des mesures. Il leur est donc possible de présenter des caryogrammes précis en longueurs développées. Ils introduisent ce faisant une représentation plus « réaliste » du caryotype, dans la mesure où elle élimine systématiquement les effets de la morphologie des chromosomes en métaphase 1404 sur leur longueur réelle. On se trouve plus proche de la constitution effective des chromosomes, d’où l’adjectif « naturelle » de l’expression « unité naturelle de longueur » intervenant dans le titre de la communication.
Mais le résultat qui leur paraît le plus important et le plus révélateur d’une propriété « naturelle » n’est pas celui-là. Avec cet instrument de mesure rendu plus stable mais aussi plus « réaliste » dirions-nous 1405 , ils peuvent soulever un autre problème et tenter de le résoudre. D’habitude, en effet, « la longueur relative de chaque type de chromosome est […] déterminée soit par rapport à un type particulièrement reconnaissable [de chromosome], soit par rapport à la longueur moyenne de l’ensemble des chromosomes de chaque cellule » 1406 . Dans ces techniques antérieures, le but, on le voit bien, consistait donc à trouver un étalon de mesure que l’on jugeait fiable soit parce qu’il était reconnaissable et donnait lieu à une mesure absolue supposée stable, soit parce qu’il était fictif mais néanmoins stable également parce que fondé sur des mesures lissées par un passage à la moyenne des mesures faites sur un même caryotype. Or, le fait expérimental que de Reffye et Essad communiquent à l’Académie prétend rendre relativement caduques de telles approches en révélant un fait naturel propre à la morphologie des chromosomes en métaphase :
‘« En effet, l’examen de ces données [les caryogrammes développés] révèle que toutes les longueurs développées obtenues pour les différents bras de chromosomes sont des multiples d’une certaine longueur que nous avons appelée ‘module’ (M). » 1407 ’Ils poursuivent :
‘« Chez le Lolium perenne, la longueur de ce module est de 0,74 μ et les bras de chromosomes en présentent de 5 à 12 […] Nous avons donc pu établir le “caryogramme modulaire“ de cette espèce. ’ ‘Il est difficile d’admettre que l’apparition de ce multiple commun aux différents bras de chromosomes soit due à un simple coïncidence. Si l’existence d’un module se confirme dans d’autres espèces, on sera conduit à considérer ce module comme un élément important de la structure et de l’évolution des chromosomes. » 1408 ’Ainsi chaque bras de chromosome est « constitué par la juxtaposition d’éléments de longueur identique et relativement importante » 1409 . Philippe de Reffye et Sadi Essad pensent avoir mis en évidence une caractéristique morphologique globale des chromosomes en métaphase. Du moins de Reffye en est-il plus convaincu qu’Essad 1410 . Ce dernier n’écarte pas en effet l’idée que cela ne soit qu’un artefact provenant des mesures. En tout cas, le caractère potentiellement significatif de ce type de travail est la raison essentielle pour laquelle Essad choisit de présenter ce résultat assez rapidement et directement à l’Académie 1411 . Grâce à une étude statistique doublée d’un modèle mathématique simple (la formule de la spirale), ils ont construit un outil de mesure qui les met en face d’une régularité morphologique autrement inaperçue. Même s’ils ne le présentent pas comme tel, le mot qu’ils proposent pour désigner cette nouvelle régularité est emprunté à la terminologie de l’architecture classique telle qu’elle apparaît dès le traité de Vitruve (1er siècle avant J.-C.) : le « module » y est l’élément architectural qui sert de mesure élémentaire à quoi l’on rapporte par proportions, par nombres entiers donc, toutes les autres mesures de l’édifice.
Autrement dit, ce qui les intéresse dans ce résultat n’est pas seulement la possibilité de mesurer la longueur réelle du chromosome mais aussi celle de décompter les nombres entiers de modules présents sur chaque bras de chromosome pour mettre au jour une architecture globale. C’est un essai de formalisation non métrique et par réitérations. Rappelons que le terme de « modèle » est un cousin très proche de celui de « module » dont il provient étymologiquement. Le module serait ici le modèle d’une construction chromosomique élémentaire au sens où sa fonction consisterait à donner lieu à une véritable architecture constituée de simples réitérations. Le module remplirait donc bien cette fonction qu’on attribue en général aux modèles et qui consiste à résumer la structure. Mais le « module » est plus réaliste que le modèle en ce qu’il prétend construire une représentation sans résidus, sans restes, sans perte d’information. À ce titre, il n’est pas une abstraction ; sa nature est bien concrète : elle est homogène à celle de l’entité biologique à laquelle on s’intéresse, en l’occurrence le chromosome. En outre, de par cette règle de construction qu’elle manifeste, la morphologie des chromosomes en métaphase est donc implicitement apparentée à un artefact humain, à un ouvrage d’ingénieur, plus précisément d’architecte. La morphologie biologique particulière du chromosome est directement apparentée à une architecture. Elle est mathématisée mais par apparentement à une science de la conception. Cependant tout en étant réaliste, car mettant au jour une régularité bien réelle présente dans le chromosome, l’hypothèse du module reste essentiellement descriptive. Les auteurs admettent ainsi qu’ils ignorent le sens ou la fonction biologique d’une telle unité chromosomique :
‘« La correspondance biologique de ce module devra être recherchée en tenant compte de la situation intermodulaire du centromère et des constrictions secondaires. » 1412 ’Autrement dit, les deux auteurs avouent ne pas pouvoir encore formuler d’hypothèse à cet égard. Comme elle ne dispose pas d’une assise fonctionnelle avérée, la suggestion du « module » n’est présentée elle-même que comme hypothèse encore à tester 1413 . La fonction du « module » dans cet article reste donc essentiellement méthodologique et descriptive. C’est tout au mieux celle d’une grille de lecture d’une organisation spatiale effective. En effet, l’application à des images de microscope d’une analyse statistique doublée d’un modèle mathématique a priori etsupposé œuvrer à la déformation des données (la formule de la spirale) a mis au jour une réalité qui n’est pas fonctionnelle : d’un point de vue biologique, il ne s’agit pas de la mise au jour d’un phénomène physico-chimique ou de régulation fonctionnelle, mais d’une entité architecturale qui, en revanche, existe bien concrètement, au niveau macro-moléculaire. En un sens, Essad et de Reffye n’ont fait que redresser des images pour les rendre plus réalistes. Le rôle du modèle mathématique est de nous permettre ici d’adopter le point de vue sur l’image recueillie le plus « réaliste » possible dès lors qu’il nous décèle cette organisation modulaire ordinairement cachée 1414 . Néanmoins, en se démarquant des approches plus fonctionnelles et physiologiques de la biologie moléculaire 1415 , les auteurs prennent la peine de prévenir une éventuelle suggestion qui pourrait leur être faite, dès lors qu’elle a de fortes chances de naître en l’esprit de tout biologiste moléculaire ou de tout cytogénéticien en ce début des années 1970 :
‘« Par ailleurs, il n’existe pas de relation directe entre ce module et le réplicon, les longueurs d’ADN impliquées dans ces deux concepts étant mesurables respectivement en millimètres et en microns. » 1416 ’Il est donc hors de question d’apparenter voire d’identifier ce module avec le réplicon 1417 qui se trouve être, en ce qui le concerne, une unité qui a été isolée pour des raisons d’abord fonctionnelles et non descriptives. Ce que veulent dire Essad et de Reffye, c’est que l’on ne peut identifier l’unité morphologique qu’ils ont mise en évidence avec l’unité fonctionnelle théorisée en biologie moléculaire. Et c’est finalement l’argument de la différence d’échelle, les millimètres par rapport aux microns, qui montre qu’une identification serait purement illusoire.
Assez paradoxalement, au premier abord, tous ces travaux originaux initiés par l’équipe d’Essad connaîtront par la suite très peu d’autres développements. Il nous est possible de trouver à cela plusieurs raisons dont la plus importante est le décès prématuré de Sadi Essad intervenu quelques années plus tard. Mais il faut bien évidemment rappeler que de son côté, de Reffye ne peut poursuivre ces travaux sur le Lolium perenne puisque, même s’il reste en contact avec Essad (qui l’encadre de loin pendant sa thèse de troisième cycle), il quitte le centre de Versailles et part, comme convenu, en Côte-d’Ivoire, juste après le DEA, afin de contribuer à l’amélioration de ces plantes arbustives bien différentes des plantes herbacées que sont les caféiers. De ce fait, ses propres travaux connaîtront inévitablement une certaine discontinuité entre le DEA et la thèse. Elle sera due, pour l’essentiel, au changement de l’objet d’étude mais aussi, comme nous le verrons, au contraste entre les différents moyens technologiques et humains mis à disposition à Versailles et en Afrique. Enfin, dans son entretien publié, Roger Cousin, qui a bien connu Essad parce qu’il était chercheur au Centre de Génétique et d’Amélioration des Plantes de Versailles depuis 1959, indique que personne à l’INRA ne voudra poursuivre exactement sur la lancée de ces travaux. Il semble cependant que Sadi Essad ait eu le temps de modifier la présentation de cette hypothèse du « module » en proposant un terme qui lui agréait davantage : celui de « nucléon » 1418 . Il ne tenait pas en effet à conserver le style un peu trop « modéliste » et théorique de l’approche que de Reffye et lui-même avaient dans un premier temps adoptée. Même sous ce nouveau terme, l’hypothèse ne sera pas non plus reprise. Enfin, dans le fait que personne à l’INRA ne poursuive ces travaux de cytogénétique au niveau chromosomique il faut sans doute voir une des conséquences de ce que les historiens des sciences Jean-Paul Gaudillière et Michel Morange ont appelé la « molécularisation » de la biologie française et plus particulièrement de la génétique formelle, entre 1965 et 1972 1419 . Les nouvelles technologies du génie génétique, appuyées notamment par le déploiement de la microscopie électronique, et intervenant pour leur part massivement au milieu des années 1970, vont également contribuer à marginaliser ce genre de travaux d’hybridation au niveau chromosomique puisque le contrôle génétique semblera de plus en plus pouvoir se faire au niveau moléculaire.
Quant à de Reffye, on le retrouve donc en poste en Côte-d’Ivoire, en ce début des années 1970. Le contexte de son institut d’accueil, l’IFCC, est bien particulier. Il est un des résultats du passé colonial de la France. Après la décolonisation, la mission de cet institut s’est résolument tournée vers le développement et vers la valorisation rationnelle des productions locales, cela dans l’intérêt de la France, bien sûr, mais aussi du pays d’accueil. En même temps, son implantation persistante dans des milieux et sous des climats bien différents de ceux de la métropole a fortement contribué à la diversification des problématiques agronomiques au-delà de ce qui aurait été possible sur le sol français. C’est dans ce contexte assez singulier donc, du point de vue de la recherche appliquée au niveau international, que prit naissance la simulation mixte.