1.2.1. Le Noyau Cochléaire (NC)
Les fibres du nerf auditif vont transmettre le message nerveux provenant de la cochlée jusqu’au noyau cochléaire. L’arrangement tonotopique observé au niveau de la cochlée se poursuit le long du nerf auditif. Les fibres apicales, codant les basses fréquences, sont localisées dans la région centrale du nerf, alors que les fibres basales, codant les hautes fréquences, sont plus localisées en périphérie (Sando, 1965).
Les fibres basales du nerf auditif vont pénétrer dans la partie dorsale du noyau cochléaire. La majorité des fibres apicales va aboutir dans le noyau cochléaire ventral, et quelques unes dans le noyau cochléaire dorsal (Lewy et Kobrak, 1936) (Figure 6).
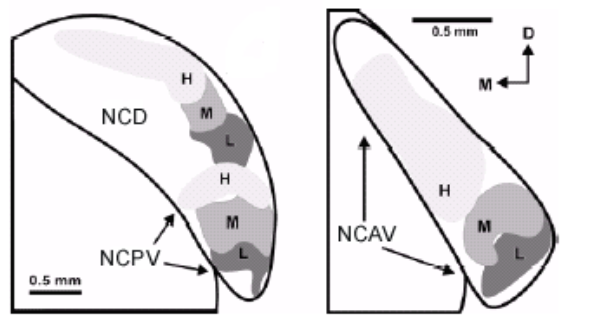
Le noyau cochléaire (NC) est le premier relais sous-cortical. Il est localisé sur la face dorso-latérale du tronc cérébral, à la jonction du pont et de la medula. Il est le premier relais auditif sur la voie ascendante et toute l’information provenant de la cochlée doit obligatoirement traverser ce noyau. Le NC peut se décomposer en 2 régions principales qui sont le noyau cochléaire dorsal (NCD) et le noyau cochléaire ventral (NCV), ce dernier étant lui-même subdivisé en noyau cochléaire antero-ventral (NCAV) et noyau cochléaire postero-ventral (NCPV).
Le rôle principal du noyau cochléaire est de traiter les informations en provenance de la cochlée, de les modifier (des systèmes d’inhibition vont permettre d’accentuer les contrastes pour la mise en forme du message sensoriel) et enfin de les distribuer vers les centres nerveux supérieurs. Pour cela, de nombreuses interconnexions existent entre les différentes parties du noyau et entre les deux noyaux cochléaires. Trois voies de sortie relaient le noyau cochléaire aux centres supérieurs : l’une vers les trois noyaux de l’olive supérieure (strie acoustique ventrale), la seconde vers les noyaux péri-olivaires (strie acoustique intermédiaire), et la dernière vers le lemnisque latéral et le colliculus inférieur controlatéral (strie acoustique dorsale dite de Monakow).
Les 3 régions principales du noyau cochléaire sont organisées de manière tonotopique. La région la plus étudiée est le NCD puisque c’est une structure bien organisée qui reçoit des afférences ordonnées du nerf auditif (Webster, 1971 ; Morest et Bohne, 1983 ; Schweitzer et Cant, 1984 ; Ryan et al, 1988).
Plusieurs techniques ont été utilisées pour révéler l’organisation fréquentielle du NCD. Des travaux basés sur l’étude du 2-deoxyglucose (2-DG) 1 chez le rat ou chez la gerbille ont montré que les neurones seraient distribués selon un gradient tonotopique. Ce dernier existe sur l’axe médiolatéral du NCD (Ryan et al., 1982 ; Ryan et Woolf, 1988 ; Ryan et al., 1988). Pour ces deux espèces, les stimuli de haute fréquence activent des régions dans la portion médiale du NCD alors que les basses fréquences activent des régions situées plus latéralement. De plus, les neurones sensibles à une fréquence donnée sont ordonnés en bandes orientées radialement sur presque toute l’épaisseur du NCD.
L’utilisation de microélectrodes implantées à l’intérieur du NCD a permis de confirmer cette organisation chez le rat (Yajima et Hayashi, 1989), chez la souris (Willott et al., 1983) et chez le chinchilla (Kaltenbach et Saunders, 1987). Les hautes fréquences caractéristiques (FC) des neurones étaient localisées à un niveau médian et les basses FC plus latéralement.
Les deux noyaux du NCV sont également organisés tonotopiquement. Cela a notamment été démontré par l’étude de l’expression du gène c-fos 2 où au niveau du NCPV et du NCAV, une augmentation de la fréquence a été corrélée à une expression de plus en plus dorsomédiane de c-fos (Saint-Marie et al., 1999a) (Figure 7).