III.1. Productions linguistique et motrice
L’influence de la motricité sur la production linguistique a principalement fait l’objet d’études comportementales ayant utilisé les techniques d’analyses de la cinématique des mouvements et du spectre vocal. Mais les études menées en TMS ont également apporté leur part de contribution en évaluant l’influence inverse de la production orale sur l’état du système moteur. Nous décrirons d’abord les résultats obtenus dans les études comportementales, nous présenterons ensuite les études de TMS.
Au niveau comportemental, les liens entre langage et motricité manuelle fine ont été évalués via la production de syllabes, unités élémentaires du discours (Gentilucci, 2003a ; Gentilucci et al., 2001, 2004a, 2004b pour une étude comparative chez les adultes et les enfants ; Gentilucci & Bernardis, 2006), mais aussi de mots (Bernardis & Gentilucci, 2006). Gentilucci et al. (2001) ont par exemple décrit une influence de l’exécution d’un mouvement de préhension sur la prononciation simultanée de syllabes. Dans leur tâche, des participants devaient saisir des objets de petite ou de grande taille avec la main droite, tout en prononçant les syllabes inscrites sur ces objets. La cinématique d’ouverture de la bouche et le spectre vocal étaient mesurés. Leurs résultats ont révélé un effet significatif de la réalisation du mouvement sur l’ouverture de la bouche (i.e. pic de vitesse d’ouverture et ouverture maximale plus grands lors de la saisie du grand objet). La taille de l’objet a également influencé le spectre vocal, à savoir que la puissance vocale maximale était plus importante si les sujets saisissaient le grand objet. Des résultats similaires ont par la suite été obtenus par Gentilucci (2003a) et Gentilucci et al. (2004ab) ayant examiné l’influence de l’observation d’actions sur la production syllabique. Dans un paradigme identique à celui précédemment décrit, Gentilucci (2003a) a ainsi rapporté une plus grande ouverture de la bouche et une émission vocale plus importante (i.e. augmentation d’amplitude des formants 6 F0 et F1) lorsque des sujets prononçaient une syllabe alors qu’ils observaient l’expérimentateur saisir un grand objet en regard d’un plus petit objet.
Ainsi, l’exécution et l’observation d’un mouvement de préhension semblent affecter à la fois l’ouverture de la bouche et le système articulatoire permettant la prononciation des syllabes, renforçant, selon les auteurs, l’hypothèse de liens fonctionnels spécifiques entre langage et motricité.Gentilucci (2003a) a notamment suggéré que l’observation du mouvement de préhension ait activé, chez les sujets, la préparation du même acte moteur, que celui-ci soit réalisé avec la main ou la bouche. Afin de conforter son hypothèse, il cite les travaux de Gentilucci et al. (2001) quant à une influence réciproque entre des mouvements réalisés par différents effecteurs distaux. Ainsi, l’exécution de mouvements de saisie avec la main peut influencer l’ouverture simultanée de la bouche, alors que la même tâche motrice réalisée avec la bouche affecte l’ouverture de la main. Selon les auteurs, la commande motrice de saisie d’objets serait envoyée aux circuits neuronaux contrôlant les mouvements des effecteurs distaux, de sorte que la réalisation d’un mouvement de la main activerait implicitement la préparation du même acte réalisé avec la bouche. Gentilucci (2003a) a alors interprété les effets observés sur la prononciation syllabique dans le cadre de cette hypothèse : l’exécution et l’observation de la saisie avec la main auraient activé un programme moteur de saisie avec la bouche. A son tour, ce programme moteur aurait influencé l’émission vocale. Autrement dit, l’exécution/observation d’actions manuelles, activant les représentations motrices de la main, aurait pu « accéder » aux représentations motrices de la bouche, impliquées dans la production verbale. Un bémol peut toutefois être apporté à cette interprétation : l’effet observé sur la production vocale pourrait n’être qu’un effet moto-moteur. Aussi, compte tenu de la proximité spatiale des représentations motrices de la main et de la bouche (Cavada, 2001 ; Matelli & Luppino, 2001 ; Rizzolatti et al., 1988), l’exécution du mouvement de préhension avec la main aurait pu « contaminer » le mouvement exécuté avec la bouche pour produire la syllabe. Une plus grande ouverture de la bouche aurait alors pu conduire à une émission vocale plus forte, effet qui ne serait pas spécifique des liens entre langage et motricité. De la même manière, une simple coactivation des motricités manuelle et orofaciale, non spécifique au langage, pourrait être à l’origine des effets obtenus dans la condition d’observation. Si l’observation du mouvement manuel a conduit à la préparation d’un acte de saisie avec la bouche, rien n’indique en effet que ce mouvement soit sélectif de l’articulation de la syllabe, et donc du langage.
En résumé, selon les auteurs (Gentilucci et al., 2001 ; Gentilucci, 2003a), l’ensemble de ces résultats supporterait « l’existence de substrats neuronaux communs aux représentations linguistiques exprimées par les gestes articulatoires et aux représentations motrices des gestes manuels », fondement de l’hypothèse d’une origine motrice du langage (Rizzolatti & Arbib, 1998). Comme nous l’avons mentionné, les éléments apportés semblent néanmoins insuffisants pour soutenir franchement une telle hypothèse.
Gentilucci et al. (2004ab) se sont ensuite intéressés à l’influence de l’exécution d’un autre type de mouvement, celui de « porter un objet à la bouche », sur la prononciation syllabique. Si le langage humain dérive d’un système de communication gestuelle, outre le mouvement de préhension, d’autres mouvements liant le bras à la bouche auraient en effet pu appartenir au répertoire d’actions de ce système. Les deux types de mouvements pourraient alors influencer sélectivement la production langagière. Afin de tester cette hypothèse, les auteurs (2004a) ont demandé à des sujets sains d’exécuter ou d’observer des mouvements de « porter un fruit à la bouche » et de prononcer la syllabe BA en fin de course du mouvement. Dans trois conditions supplémentaires, soit les sujets émettaient une vocalisation n’existant pas dans la langue italienne (OE) en lieu et place de la syllabe, soit ils observaient l’expérimentateur mimer l’action avec la main ou à l’aide d’un effecteur non biologique (en absence de fruit ; l’expérimentateur n’ouvrait pas la bouche). Les mouvements étaient réalisés sur une cerise (objet de petite taille) ou une pomme (objet de grande taille). D’une part, les résultats ont révélé que l’exécution et l’observation des mouvements transitifs et intransitifs (mime) ont affecté l’émission vocale (i.e. augmentation d’amplitude du formant F2 lorsque l’objet porté à la bouche était la pomme). L’obtention du même pattern de résultats, que l’action observée soit réalisée sur le fruit ou simplement mimée, a conduit les auteurs à écarter l’éventualité d’un effet purement lié à l’observation du fruit ou de l’ouverture de la bouche de l’expérimentateur. Elle est également congruente avec les études ayant décrit l’existence d’un système miroir impliqué dans la reconnaissance des actions transitives mais aussi intransitives chez l’homme (Buccino et al., 2001, 2004a ; Fadiga et al., 1995 ; Maeda et al., 2002). D’autre part, un effet a été obtenu sur la cinématique d’ouverture de la bouche (i.e. latence et amplitude du pic d’ouverture plus grandes lors de la saisie de la pomme) uniquement lorsque les mouvements observés étaient transitifs, c’est-à-dire lorsque l’expérimentateur ouvrait la bouche pour croquer le fruit (en accord avec Gentilucci et al., 2001, et Gentilucci, 2003a). Cet effet ne pourrait-il toutefois simplement résulter de l’imitation de l’ouverture de la bouche par les sujets ? Enfin, aucun effet sur le spectre vocal n’a été retrouvé lorsque les participants devaient produire la vocalisation (OE) ou lorsque le mime du mouvement observé était réalisé avec l’effecteur non biologique. Ces données semblent donc conforter l’idée (i) de liens spécifiques entre système linguistique et motricité et (ii) que le stimulus biologique en mouvement soit un pré-requis nécessaire à l’émergence des liens entre langage et motricité (Gentilucci, 2003a). Conformément à leurs précédentes interprétations, les auteurs ont suggéré que l’exécution et l’observation du mouvement manuel de « porter un fruit à la bouche » aient implicitement activé la préparation du mouvement de « manipuler de la nourriture avec la bouche » (i.e. mâcher). La représentation motrice de cet acte aurait alors influencé l’ouverture de la bouche mais aussi l’émission de la voyelle « a » (F2) lors de la prononciation de la syllabe. La même critique d’un effet moto-moteur de la main à la bouche, plutôt que spécifique des liens entre langage et motricité, peut là encore s’appliquer à cette interprétation. Les effets obtenus uniquement sur le spectre vocal, et non sur la cinématique d’ouverture de la bouche, lors de l’observation de mouvements intransitifs pourraient quant à eux suggérer une influence directe de la motricité manuelle sur la motricité du larynx pour la production vocale.
Pour Gentilucci et al. (2004a), l’hypothèse formulée permettrait également de rendre compte des effets distincts de l’exécution/observation du mouvement de « porter un fruit à la bouche » et du mouvement de préhension manuelle sur la production syllabique (voir aussi Gentilucci et al., 2004b). En effet, alors que le premier mouvement a principalement affecté le formant F2, le deuxième a surtout influencé F1. Les deux types d’actions semblent ainsi induire des changements distincts d’articulation et de positionnement de la bouche, et par conséquent, influencent la production vocale de manière différente. En d’autres termes, « les conséquences motrices des actions manuelles seraient transcrites en postures articulatoires spécifiques affectant les formants ». Gentilucci et al. (2004b) ont même proposé que certaines voyelles puissent dériver de différentes actions manuelles. Selon cette hypothèse, certains phonèmes découleraient de postures articulatoires de la bouche induites par des gestes manuels particuliers : ces phonèmes auraient émergé pour exprimer le but d’une action manuelle. Ces sons liés aux actions auraient été initialement utilisés pour communiquer entre individus de la même espèce (e.g. le son associé au mouvement de préhension aurait eu le sens de « prendre possession de », alors que celui associé au geste de porter à la bouche aurait signifié « objet comestible »). Puis, les interactions sociales se complexifiant, les hommes auraient ajouté d’autres phonèmes à ces sons afin de construire des mots plus élaborés et d’en préciser le sens. Certains aspects du sens original de ces phonèmes persisteraient toutefois, rendant compte, pour les auteurs, des effets de la motricité manuelle sur la production syllabique. L’hypothèse avancée par Gentilucci et collègues (2004b) ne semble toutefois reposer que sur de pures spéculations et paraît difficile à corroborer au vu des données disponibles actuellement.
L’ensemble des résultats comportementaux présentés démontre donc que l’exécution, et même l’observation d’actions manuelles, qu’elles soient transitives ou non, peuvent influencer la production syllabique. Si les auteurs suggèrent que ces données s’accordent clairement avec l’hypothèse d’un système primitif « miroir » basé sur la communication brachio-manuelle comme précurseur du langage (Corballis, 2002 ; Rizzolatti & Arbib, 1998), les effets observés pourraient néanmoins émerger comme conséquence d’une co-activation des motricités manuelle et orofaciale, cette dernière n’étant pas nécessairement spécifique aux gestes articulatoires et donc au langage.
Comme nous l’avons mentionné plus haut, les études menées en TMS se sont intéressées à l’influence de la production verbale sur l’excitabilité du système moteur (Floël et al., 2003 ; Meister et al., 2003 ; Tokimura et al., 1996). Tokimura et ses collègues (1996) ont par exemple décrit une augmentation d’amplitude des potentiels évoqués moteurs (PEMs) enregistrés sur les muscles de la main droite (stimulation hémisphérique gauche) lors de la lecture de textes à voix haute. Aucun effet facilitateur n’a été observé lorsque la stimulation était délivrée dans l’hémisphère droit non dominant du langage (PEMs sur la main gauche). Pour la première fois, des effets latéralisés de la production linguistique sur le système moteur manuel ont donc été mis en évidence (mais voir Floël et al., 2003 pour des effets bilatéraux). Plus récemment, Meister et ses collègues (2003) ont demandé à des sujets sains de lire à voix haute des noms concrets isolés ou en triplets (non liés sémantiquement), alors que les aires motrices de la main et de la jambe de leurs deux hémisphères étaient stimulées. L’utilisation de stimuli de différentes longueurs leur a ainsi permis de déterminer si l’augmentation potentielle d’excitabilité motrice lors de la lecture n’était induite qu’au début de la production ou persistait pendant toute la durée de l’articulation. En outre, la TMS était délivrée à différents intervalles, soit avant, pendant ou après la tâche verbale (Figure 3.1.a pour la séquence temporelle des stimuli). Conformément aux résultats de Tokimura et al. (1996), ils ont démontré une augmentation de l’amplitude des PEMs enregistrés sur les muscles de la main droite lorsque la stimulation était appliquée sur le cortex moteur gauche pendant la lecture (Figure 3.1.b). Aucun effet n’est apparu pour les stimulations précédant et suivant la tâche verbale, ni pour celles appliquées sur l’hémisphère droit. Enfin, la tâche verbale n’affectait pas l’amplitude des PEMs enregistrés sur les muscles de la jambe (i.e. stimulation de l’aire motrice de la jambe), soulignant le rôle de la motricité manuelle dans le traitement du langage. Ces résultats confirment donc que la production linguistique peut affecter l’excitabilité corticale motrice, les effets étant restreints à l’hémisphère gauche. En outre, une corrélation entre les effets facilitateurs et la durée de la tâche verbale a été mise en évidence, suggérant des liens fonctionnels spécifiques entre langage et système moteur. Ainsi, l’augmentation des PEMs enregistrés sur la main droite durait tant que la tâche verbale était en cours (Figure 3.1.b).
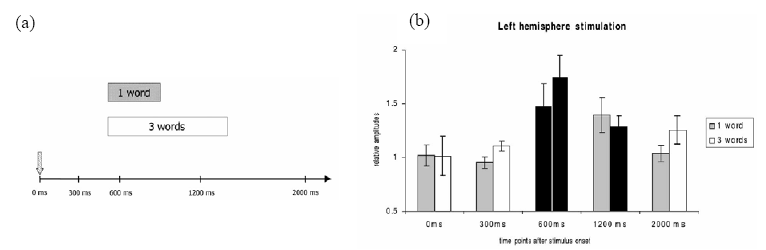
(a) Séquence temporelle des stimuli. La flèche indique le moment de présentation des stimuli (mots isolés ou triplets de mots). En moyenne, les temps de réponse des sujets étaient de 500 ms (i.e. la production orale débutait au temps t = 500 ms). La durée moyenne de prononciation des mots isolés était de 400 ms (barre horizontale grise) et celle des triplets de mots de 1000 ms (barre horizontale blanche). Les stimulations magnétiques étaient délivrées à différents intervalles après la présentation des mots : 0, 300, 600, 1200 et 2000 ms. (b) Résultats. Amplitude moyenne des PEMs enregistrés sur les muscles de la main droite, lors de la stimulation du cortex moteur gauche. Les résultats sont présentés pour les différents temps de stimulation magnétique. Les barres noires indiquent les conditions significativement différentes de la ligne de base. L’effet facilitateur était observé lors d’une stimulation à 600 ms, soit 100 ms après le début de la production orale, pour les mots isolés et en triplets. Pour les triplets, il était également obtenu pour une stimulation à 1200 ms (la tâche de production de ces triplets était encore en cours alors que celle des mots isolés était terminée). Pris de Meister et al. (2003).
Ces études menées en TMS démontrent donc clairement un recrutement de l’aire motrice gauche de la main lors de la production linguistique, témoin probable des connexions étroites entre les centres du langage et le système moteur manuel établies lors du développement du langage. De la même manière que pour les études comportementales, un effet moto-moteur pourrait néanmoins être partiellement à l’origine des résultats obtenus. En raison de la proximité spatiale des représentations motrices de la main et de la bouche, la réalisation de mouvements avec la bouche lors de la lecture à voix haute pourrait affecter l’excitabilité motrice de l’aire de la main sans que cette influence soit spécifique au langage. Le fait que les effets soient latéralisés dans l’hémisphère gauche, dominant du langage, conforte toutefois l’hypothèse de liens intimes entre production verbale et motricité manuelle.
Comme nous l’avons décrit au travers de la revue des différentes études menées jusqu’à présent, la production linguistique semble activer des aires corticales initialement dévolues à la motricité manuelle, mais qui, au cours de l’évolution, auraient été recrutées dans un but communicatif, devenant ainsi les aires « précurseurs » du langage humain. Qu’en est-il alors de la perception du langage ? Si comme le postule la théorie motrice de la perception du langage, les processus de production et de perception linguistiques partagent des substrats neuronaux communs, la perception de mots devrait influencer l’excitabilité corticale motrice de la même manière. Nous allons maintenant présenter les travaux, menés en TMS, ayant examiné cette hypothèse.