Chapitre I. Introduction générale
‘«What if we awake one day, all of us, and find ourselves utterly unable to read?I wish you to gasp not only at what you read but at the miracle of its being readable. »
Vladimir NABOKOV, pale fire (1962)’
I.1. Le langage humain
Une langue, par définition, est un système d'expression et de communication commun à un groupe social (communauté linguistique) (Le Robert méthodique, 1982). Elle est la manifestation et la concrétisation du langage, particulière à une certaine communauté, alors que le langage est une faculté inhérente, universelle, et spécifique à l’espèce humaine. Si la communication n’est pas le propre de l’homme, le langage humain diffère cependant de la communication animale dans le sens où il permet d’exprimer un nombre infini d’idées et de produire des affirmations, des requêtes, etc. Cette spécificité du langage humain tient à sa structure, basée sur une double combinatoire : les sons sont combinés pour former des mots, les mots sont combinés pour former des phrases (Dortier, 2001). Alors, quand et comment est apparu le langage?
L’évolution des capacités mentales caractéristiques de l’espèce humaine est couramment considérée comme corrélée à la naissance de l’Homo erectus, il y a quelques deux millions d’années. Depuis, un accroissement rapide de la taille du cortex, surtout frontal et temporal, se serait produit (Wood and Collard, 1999). De tels changements permettraient aux individus de mettre en place des fonctions cognitives telles que la théorie de l’esprit et l’imagerie mentale, qui sont nécessaires pour la coopération sociale, la planification d’action et le langage (Dortier, 2001; Corballis, 2005). Toutefois, tandis que pour certains chercheurs la protolangue (une forme d’expression consistant à assembler quelques mots dans un énoncé court, sans support grammatical ; Bickerton, 1990) serait apparu avec l’Homo erectus, pour d’autres la langue n’est pas apparue avant la naissance de l’homme moderne (Homo sapiens sapiens) voici quelques 170 000 ans (Ingman et al., 2000; Corballis, 2005).
Si l’origine du langage est une question fondamentale posée aux sciences du langage, il n’est pas évident d’y répondre, surtout parce que le langage est un comportement, et qu’on n’en trouve donc pas de trace directe dans les vestiges du lointain passé des espèces qui ont précédé l’espèce humaine. Grossièrement, on peut diviser les théories de l’évolution du langage en deux grands courants: celles qui considèrent le langage comme le produit d’un processus propre d’adaptation, et celles qui présentent le langage comme une conséquence de l’évolution d’autres capacités. (voir Reboul, 2007)
Le premier courant est connu sous le nom de courant adaptationniste. Le terme adaptation dérive de la théorie de l’évolution proposée par Darwin. L’idée est qu’un caractère existe dans une population parce qu’il est sélectionné par l’environnement en raison de l’avantage adaptatif qu’il confère à l’individu qui le porte (Darwin, 1859). Il faut noter que cette conception ne concerne pas l’acquisition de certaines propriétés au cours de l’ontogénie (au niveau de l’individu), mais consiste seulement en un processus qui sélectionne une variation favorable, dont l’apparition reste le fait du hasard. Pinker (eg. Pinker and Bloom, 1990), par exemple, adopte cette position et propose que le langage possède un « design » compliqué et que la seule explication pour sa complexité est la sélection naturelle. Pourtant, pour appliquer une explication adaptationniste à l’évolution du langage, il faut indiquer des raisons (environnement) pour lesquelles le langage a été sélectionné. L’hypothèse sociale en est une, proposant que le langage ait évolué pour l’avantage « social » qu’il procure (Dunbar, 1996). Selon les défenseurs de l’hypothèse sociale, le langage est sélectionné par la nécessité de résoudre les nombreux problèmes que pose une vie sociale riche et complexe. Vu que le mot « social » a été utilisé par les différents auteurs dans des sens différents, une version alternative de cette hypothèse, l’hypothèse mixte cognitivo-sociale, a été proposée (Newmeyer, 2003). Cette dernière est une hypothèse plus précise et plus couramment admise de nos jours. Elle passe du « social » comme « maintenir la cohésion du groupe » au « social » comme signifiant « communiquer des informations ». Selon cette hypothèse, le langage a évolué pour permettre la communication d’informations qui est considérée comme une fonction cognitive autant qu’une fonction sociale.
Le deuxième courant, dit aussi le courant exaptationniste, suppose que le langage n’est qu’un sous-produit d’autres évolutions. La notion d’exaptation a été introduite par Gould et al. (Gould, 1980). L’idée est que parmi les traits biologiquement déterminés, si certains sont des adaptations, une grande partie est due au hasard – ce ne sont pas les fruits d’un processus d’adaptions, mais des sous-produits : des pré-conditions d’un trait adaptatif ou des conséquences non-sélectionnées d’une authentique adaptation. Chomsky (Chomsky, 1966; Chomsky, 1986), par exemple, pense que le système biologique (i.e. inné) sous-tendant le langage n’est qu’un sous-produit d’autres adaptations. La théorie exaptationniste culturelle (Tomasello, 1999) est une autre théorie importante dans ce courant. Elle propose que le langage est le fruit d’un processus d’évolution culturelle (i.e. acquis) et non pas d’un processus d’évolution biologique, étant donné que l’évolution culturelle va plus vite au niveau de la transmission (d’un membre à d’autres membres dans un groupe) et est cumulative (on peut s’attendre à une évolution des comportements au cours du temps). Il faut noter que, selon cette théorie, la langue n’est pas elle-même une exaptation mais un produit (culturel) d’une exaptation, alors que l’évolution culturelle est une exaptation, rendue possible par l’adaptation biologique qui serait la capacité à interpréter le comportement d’autrui (cf. Reboul, 2007).Une autre hypothèse dans ce courant, l’hypothèse du protolangage proposé par Bickerton (1990) est admise plus couramment. Cette hypothèse propose que nos capacités linguistiques aient évolué en deux étapes : d’abord, l’Homo erectus aurait communiqué au moyen d’une protolangue voici deux millions d’années; puis le protolangage se serait enrichi d’une syntaxe plus formelle permettant d’affiner la précision et la clarté de nos échanges, voici environ 50 000 ans. Il suppose que l’apparition de cette compétence syntaxique s’est produite suite à une exaptation (cf. Calvin and Bickerton, 2000)
Par ailleurs, certains chercheurs ne font pas partie de ces deux voies – ils ne supposent pas du tout que le langage a évolué (Chomsky, 2006). Tandis que précédemment la position de Chomsky était plutôt pour la théorie exaptationniste (« sous-produit »), dans des articles récents, il fait l’hypothèse que le langage ne serait pas nécessairement indépendant des autres processus cognitifs et que sa spécificité pourrait être très réduite et être le fruit d’un processus auto-organisationnel quelconque. Ceci soulève cependant, une question cruciale : pourquoi le langage existe-t-il dans l’espèce humaine et seulement dans l’espèce humaine? (cf. Reboul, 2007).
En dépit d’une littérature florissante, les études sur l’origine du langage, reposant surtout sur les découvertes archéologiques, ne semblent donc que peu concluantes. A cet égard, les études génétiques offrent une perspective intéressante, bien que les découvertes soient encore fragmentaires.
L’homme et le chimpanzé (notre voisin le plus proche) partagent 99% de leur patrimoine génétique (Ebersberger et al., 2002). Si la culture doit jouer un certain rôle dans l’évolution du langage, elle n’explique pas l’incapacité des chimpanzés, élevés dans un environnement humain, à acquérir une réelle compétence linguistique (Terrace et al., 1979). En 2001, des chercheurs ont identifié un premier gène candidat de l’évolution du langage – FOXP2 (Lai et al., 2001). Au sein d’une famille anglaise connue sous le nom de KE, certains membres présentaient un étrange trouble: non seulement ils avaient des problèmes de grammaire, d’écriture et de compréhension, mais ils avaient aussi un déficit développemental d’acquisition de la parole, caractérisé par des troubles sévères d’articulation (i.e. dyspraxie oro-faciale, c’est-à-dire l’incapacité à coordonner les mouvements du visage, de la bouche et de la langue pour articuler correctement les mots ou les non-mots multi-syllabiques) (Gopnik and Crago, 1991; Vargha-Khadem et al., 1995, 1998). Un autre individu, hors de la famille KE, a un trouble de l’articulation similaire à celle de la famille KE. Les chercheurs ont trouvé chez ces patients une version mutée du gène FOXP2, situé sur le chromosome 7 (7q31 ; Lai et al., 2001). Ils en ont donc déduit que ce gène, dans sa version humaine, était essentiel à l'apparition d'un langage articulé.
Si des variantes de ce gène sont identifiées chez toutes les espèces de mammifères, la protéine FOXP2 chez l’homme diverge par trois acides aminés (sur 751 au total) par rapport à la version présente chez la souris, et deux par rapport à la version chez le chimpanzé. Cela indique donc une pression sélective récente et forte: l’ancêtre commun des primates et des rongeurs a vécu voici 130 millions d’années (un acide aminé modifié), alors que celui des chimpanzés et des hommes vivait il y a seulement 6 millions d’années (deux acides aminés modifiés) (Enard et al., 2002). Une étude récente (Krause et al., 2007) a identifié, chez l’homme de Néandertal (Homo sapiens neanderthalensis), la même version du gène FOXP2 que celui chez l’homme. Les auteurs proposent aussi que le balayage sélectif (le processus qu’une mutation avantageuse remplace toute autre variation présente à ce locus) se soit passé voici quelques 300 000 ans, avant la séparation de ces deux espèces.
L’analyse des modèles animaux suggère que le gène FoxP21 joue d’abord un rôle dans l’apprentissage et la coordination des mouvements complexes, d’où son influence sur le langage humain à travers l’articulation des mots (Teramitsu and White, 2008). Particulièrement, puisque la parole chez l’homme et les vocalisations acquises chez les oiseaux chanteurs présentent des parallèles comportementaux et neuraux, les oiseaux chanteurs fournissent un excellent modèle pour étudier le mécanisme sous-tendant la parole chez l’homme. Des études par le groupe de Scharff ont montré que le gène FoxP2 joue un rôle important dans l’apprentissage moteur vocal auditivement guidé (dans les ganglions de la base) chez les oiseaux chanteurs comme chez l’homme (Rochefort et al., 2007), et le « knockdown » du gène FoxP2 (i.e. éteindre/‘réduire au silence’ systématiquement l'expression de ce gène en utilisant le technique d’ANR interférence) chez les oiseux chanteurs aboutit à une imitation incomplète et inexacte des chansons du tuteur (Haesler et al., 2007).Ces résultats suggèrent par ailleurs une connexion dans le cerveau entre les régions sous-tendant la perception auditive et les régions sous-tendant le mouvement vocal chez les oiseaux chanteurs comme chez l’homme.
Néanmoins, le gène FOXP2 s’exprime en fait non seulement dans le cerveau (le cervelet, les ganglions de la base et les régions frontales inférieures) mais aussi dans d’autres tissus tels que les poumons, l’œsophage et le cœur. Les travaux préliminaires du groupe de Daniel H. Geschwind ont récemment montré que le gène FOXP2, comme plusieurs autres gènes, serait impliqué dans le mécanisme de la croissance des neurones et de leur connectivité dès le début du développement nerveux fœtal (Vernes et al., 2007). Ainsi, bien que le gène FOXP2 ait une fonction importante dans le langage, il ne peut pas être considéré comme le gène du langage humain : il semble être impliqué dans un mécanisme comprenant le langage mais d’une manière plus générale, tandis que d’autres gènes impliqués dans le langage resteraient encore à découvrir.
Si l’on regarde les différences au niveau du cerveau entre les espèces de primates humains et non-humains, elle ne concerne pas seulement la taille, mais aussi la structure cérébrale. Une étude très récente (Rilling et al., 2008) a montré une grande différence de faisceau arqué (un ensemble d'axones impliqué dans le langage humain) entre l’homme, le chimpanzé et le macaque. Rilling et al. (2008) ont observé, en utilisant la méthode de tractographie, une projection saillante du faisceau arqué vers le lobe temporal chez l’homme tandis que cette projection est absente ou très faible chez les primates non-humains (Figure I.1). Selon les auteurs, la spécialisation de cette projection est probablement liée à l’évolution de langage.
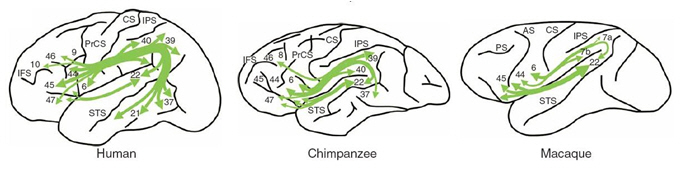
Certes, ces informations disponibles restent fragmentaires, pourtant les progrès accomplis dans les différents domaines nous permettent de faire avancer la discussion et d’approcher la vérité de l’évolution de langage. Ce qui est certain est que notre capacité du langage repose sur un cerveau équipé de sa propre structure avec des régions cérébrales interconnectées ; ce cerveau rend possible notre capacité ultérieure à apprendre à écrire et à lire.